
Biogenesis of Melanosomes in Melanocytes
- 演化之聲

- May 29
- 5 min read
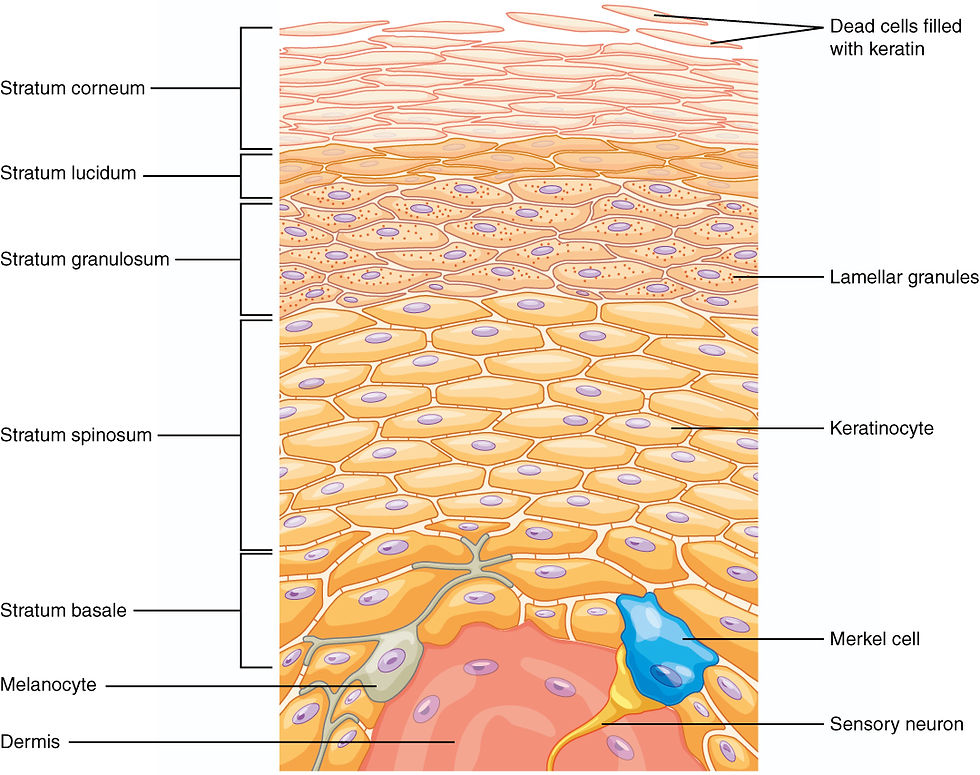
Melanocytes are derived from embryonic neural crest cells. After closure of the neural tube, a population of immature melanoblasts begins to migrate to sites including the basal layer of the epidermis, or stratum basale, and the hair follicles. Their migration, settlement, and maturation are regulated by multiple signaling factors. For example, stem cell factor (SCF) binds to the c-Kit receptor on the surface of melanoblasts and melanocytes, inducing their proliferation and differentiation into mature melanocytes capable of producing melanin. Defects in these genes can lead to the congenital pigmentation disorder piebaldism. Once melanocytes have reached the epidermis, they extend slender dendrites within the basal layer and establish connections with approximately 30 to 40 surrounding keratinocytes.




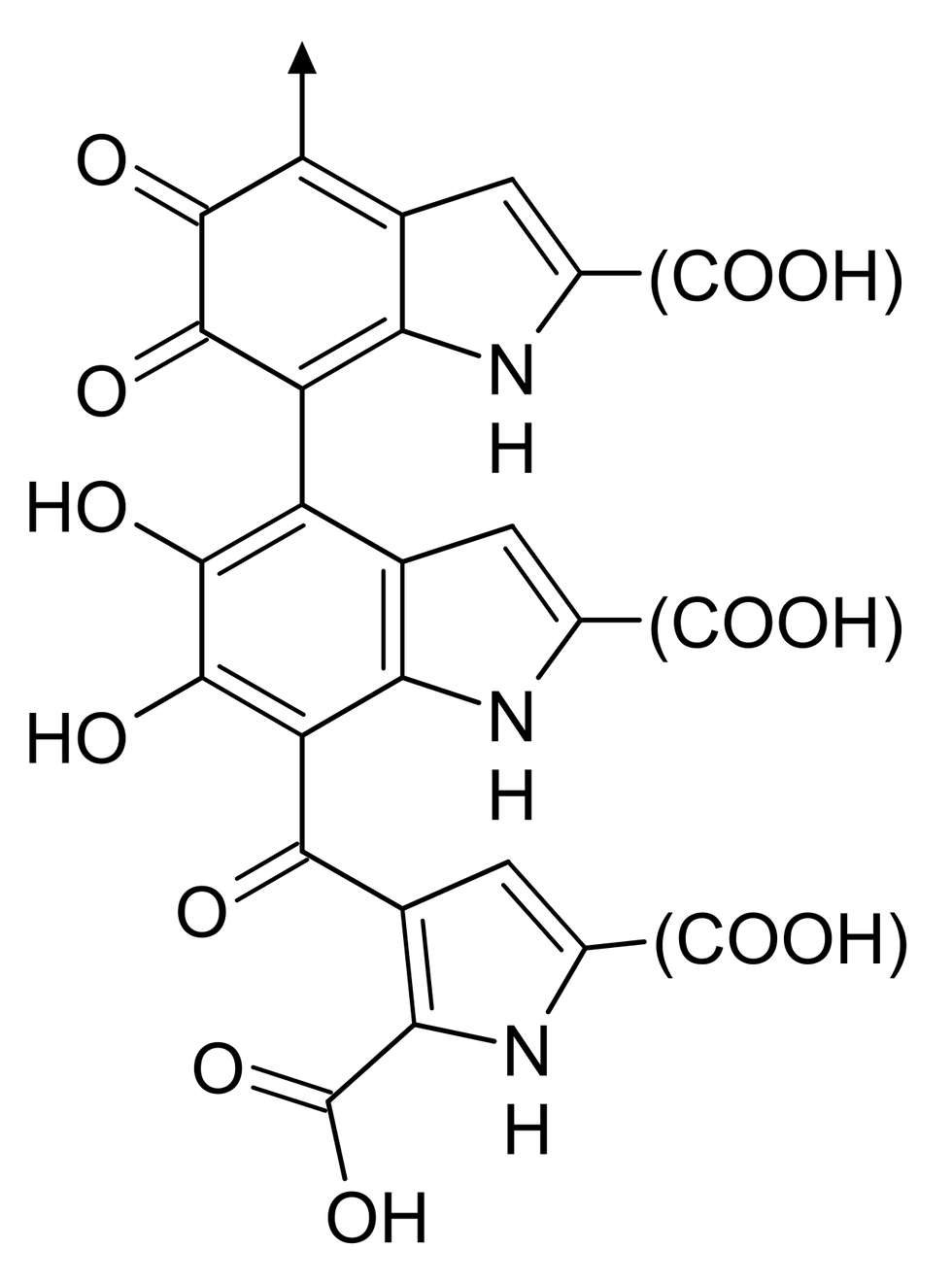
In humans, melanin can be divided into two major classes: black or brown eumelanin and yellow or red pheomelanin. Eumelanin is derived from 5,6-dihydroxyindole (DHI) or 5,6-dihydroxyindole-2-carboxylic acid (DHICA), whereas pheomelanin is derived from benzothiazine or benzothiazole. Melanin is synthesized and stored within melanosomes inside melanocytes. Melanosomes are lysosome-related organelles that share certain structural and molecular features with lysosomes, while possessing the specialized ability to synthesize and carry melanin.

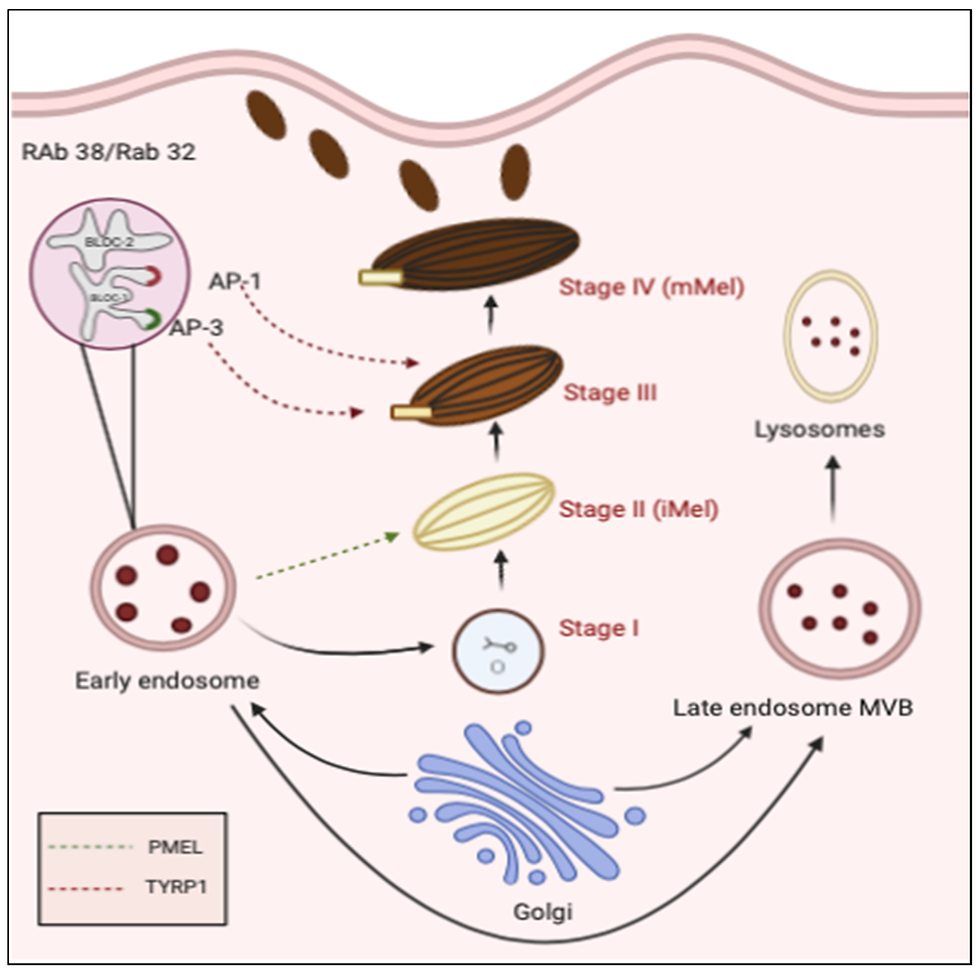
Melanosome maturation proceeds through four stages. Stage I melanosomes are relatively round vesicles with amorphous contents, in which intraluminal fibrils begin to form. By stage II, these fibrils have become organized into a distinct internal meshwork. Because substantial melanin deposition has not yet occurred, stages I and II are commonly regarded as early melanosomes. The principal structural protein of this internal framework is PMEL. Following its processing and trafficking into early melanosomes, PMEL forms a fibrillar matrix that subsequently binds melanin, allowing the pigment to be stably deposited within the organelle. At stage III, melanin synthesis becomes active and pigment is progressively deposited on the fibrillar matrix, gradually obscuring the previously distinct internal structure. By stage IV, the melanosome is highly mature and densely filled with melanin, so that the original arrangement of intraluminal fibrils is almost no longer visible. At this point, mature melanosomes are ready to be transferred to keratinocytes.
Tyrosinase initiates the early steps of melanin synthesis. It is a copper-dependent membrane-bound enzyme that converts L-tyrosine into L-DOPA, thereby driving the subsequent reactions involved in the synthesis of different forms of melanin. If tyrosinase loses its function, melanin cannot be produced normally, resulting in the severe pigmentation disorder oculocutaneous albinism type 1. Melanosomes also contain tyrosinase-related protein 1 (TYRP1) and dopachrome tautomerase (DCT, also known as TYRP2), which participate in melanin formation and in the regulation of melanin type.

Newly formed melanosomes are generally located near the nucleus. As they mature, they move toward the cell periphery through a transport system involving microtubules, kinesin, and dynein. When melanosomes approach the tips of the dendrites, the F-actin-based transport system takes over. Through a complex formed by Rab27a, melanophilin, and myosin Va, melanosomes are positioned near the plasma membrane. Mature melanosomes are then transferred to keratinocytes. This transfer process has not yet been fully clarified, and proposed mechanisms include exocytosis followed by endocytosis, cytophagocytosis of melanocyte dendrite tips by keratinocytes, fusion of plasma membranes, and transport through membrane vesicles. After entering keratinocytes, melanosomes gather around the nucleus, with most of them positioned above the nucleus on the side facing the skin surface. This cap-like arrangement shields the nucleus from incoming ultraviolet light, reducing UV-induced damage to the DNA within it.


Ultraviolet light from sunlight can darken the skin through responses that occur over two different timescales. Ultraviolet radiation can be broadly divided into three types: long-wave ultraviolet A (UVA, 315-400 nm), medium-wave ultraviolet B (UVB, 280-315 nm), and short-wave ultraviolet C (UVC, 100-280 nm). UVA has the lowest energy of the three but the greatest penetrating ability, allowing it to reach the dermis. UVB has higher energy than UVA, acts primarily on the epidermis, and readily causes direct DNA damage. UVC has the highest energy and an exceptionally strong capacity to damage DNA, but it is absorbed almost completely by the ozone layer and therefore ordinarily does not reach the Earth's surface. The immediate pigment darkening induced by UVA wavelengths can appear during exposure or shortly afterward and generally persists for several hours to approximately one day. During this response, the darkening of the skin results primarily from oxidation of existing melanin in melanocytes and keratinocytes after UVA stimulation, together with redistribution and movement of pre-existing melanosomes. There is no substantial increase in new melanin synthesis, and the photoprotective capacity of this response is limited.
Delayed tanning is the response most closely associated with newly synthesized melanin. It begins several days after sufficient ultraviolet exposure and may persist for weeks or even longer. This response is driven largely by UVB because its principal mechanism is initiated by DNA damage. Its characteristic features include increased melanin synthesis, increased melanosome number and melanization, greater dendricity of melanocytes, and enhanced transfer of melanin to keratinocytes. Delayed tanning begins in keratinocytes. When UVB causes DNA damage in these cells, the tumor suppressor protein p53 is activated, promoting expression of the pro-opiomelanocortin (POMC) gene. After POMC is cleaved, it gives rise to multiple peptides, including α-melanocyte-stimulating hormone (α-MSH), adrenocorticotropic hormone (ACTH), and β-endorphin. α-MSH and ACTH can act on the MC1R receptor on the melanocyte membrane, increasing intracellular cyclic AMP (cAMP). Through protein kinase A and CREB-associated transcriptional regulation, this signaling ultimately increases expression of the melanocyte-specific MITF transcription factor. MITF then promotes the expression of proteins including tyrosinase, TYRP1, DCT, and PMEL, enabling melanocytes to produce a greater number of more mature melanosomes and transfer them to keratinocytes.

The skin's response to ultraviolet light, however, is not mediated solely through the α-MSH pathway; the pathway described above represents only one component of a broader stress-response network activated cooperatively by multiple cell types within the epidermis. Darkening of the skin is therefore a defensive response initiated when the skin is subjected to photic stress. Melanin itself cannot remove DNA damage that ultraviolet light has already caused; that task belongs to the DNA repair systems. What melanin can provide is an additional natural, although limited, layer of protection during future ultraviolet exposure.
Author: Shui-Ye You
References:
Bao M et al. (2025). Melanosome Transport and Processing in Skin Pigmentation: Mechanisms and Targets for Pigmentation Modulation. Int. J. Mol. Sci.
dos Santos Videira IF et al. (2013). Mechanisms regulating melanogenesis. An Bras Dermatol.
Gabbianelli M and Testa U. (2009). Role Of Stem Cell Factor In The Reactivation Of Human Fetal Hemoglobin. Mediterr J Hematol Infect Dis.
Gilchrest BA et al. (1996). Mechanisms of ultraviolet light-induced pigmentation. Photochem Photobiol.
Khan AQ et al. (2020). Exosomes: Emerging Diagnostic and Therapeutic Targets in Cutaneous Diseases. Int. J. Mol. Sci.
Robinson GN and Pickering RJ. (2024). Melanocortins and Their Potential for the Treatment, Prevention and Amelioration of Complications of Diabetes. Diabetology.
Snyman M et al. (2024). The metabolism of melanin synthesis—From melanocytes to melanoma. Pigment Cell & Melanoma Research.
Yamaguchi Y and Hearing VJ. (2009). Physiological factors that regulate skin pigmentation. Biofactors.
Yardman-Frank JM and Fisher DE. (2021). Skin pigmentation and its control: From ultraviolet radiation to stem cells. Exp Dermatol.



Comments