
Does Lipoptena andaluciensis still need vision after it shifts from the winged form to an ectoparasitic life?
- 演化之聲

- Jun 16
- 4 min read
The deer ked Lipoptena andaluciensis still has wings when it first emerges as an adult. At this stage, it leaves its birthplace and flies through woodland edges and clearings in search of large mammalian hosts, such as roe deer (Capreolus capreolus), fallow deer (Dama dama) and red deer (Cervus elaphus). During this host-seeking phase, it must detect distant objects, avoid obstacles and use differences in colour and brightness to approach a suitable host. Once it finds one, however, its life changes dramatically: it breaks off its own wings, moves into the host’s fur and becomes an ectoparasite. From that point onward, it no longer needs to fly. Its priorities shift toward blood feeding, digestion, mating and reproduction.
This raises an intriguing question: when an insect permanently foregoes flight, does it also abandon its energetically costly visual system? Sensory systems are essential for animal behaviour, but they are expensive to maintain. Retinas, photoreceptive proteins and neural processing all require energy. If vision becomes less useful during the ectoparasitic stage, natural selection would be expected to favour reduced investment in unnecessary visual machinery. L. andaluciensis is a particularly interesting species for testing this idea because it passes from an alate, host-seeking adult stage to a dealate, ectoparasitic adult stage without moulting or undergoing another metamorphosis. It cannot grow a new kind of eye after settling on a host. If its visual function declines, the change must occur within the same visual system.
The researchers examined the head of L. andaluciensis using scanning electron microscopy. The head is wider than it is long and is dorsoventrally flattened. Three ocelli are present in a triangular arrangement on the posterior part of the dorsal surface. The compound eyes lie at the anterolateral margins of the head and wrap around toward the lateral, dorsal and ventral surfaces. Each compound eye has roughly 500–550 facets, fewer than those of Drosophila, Musca and Calliphora. The facets are not uniform in shape: they are more nearly hexagonal anteromedially, but become more square laterally and dorsally. These external head structures do not appear to change substantially between alate and dealate adults.
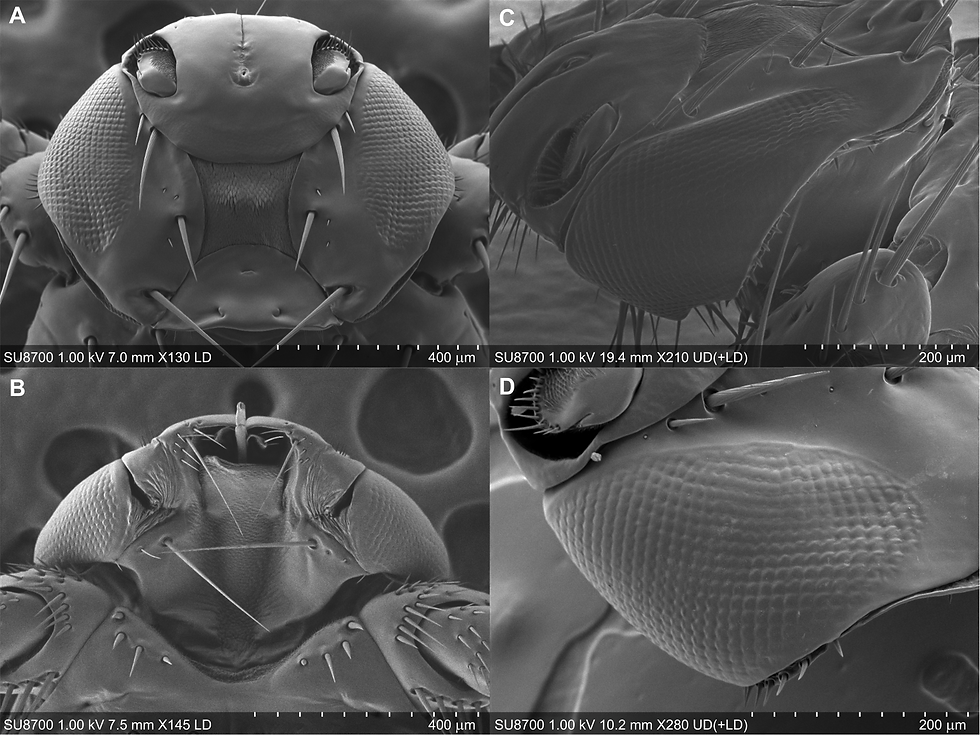
The researchers then compared the transcriptomes of alate and dealate L. andaluciensis. The alate, host-seeking adults were collected along woodland edges and clearings in the Tuscan Apennines of Italy, whereas the dealate ectoparasitic adults were collected from the carcasses of legally hunted red deer. The transcriptome data showed that L. andaluciensis retains five opsin types typical of the visual system of higher dipteran flies. Rh1 is expressed by the outer photoreceptors R1–R6 of the compound eye and mainly supports motion vision, allowing the fly to detect object movement and changes in the visual field during flight. Rh3, Rh5 and Rh6 are associated with ultraviolet-, blue- and green-sensitive photoreceptors, forming the basis of colour vision. Rh2 is expressed in the ocelli, which contribute to stabilisation reflexes and are especially important for flight control.
The transcriptomic analysis showed that these opsins were still expressed in the heads of dealate ectoparasitic adults. They had not disappeared. In other words, after breaking off their wings, the flies did not shut down the molecular machinery for colour vision, motion vision or ocellar photoreception. However, opsin expression was clearly reduced, falling to roughly half of the level seen in alate host-seeking adults. This provides a careful answer to the central question: after L. andaluciensis foregoes flight, its visual system is not removed, but opsin gene expression is downregulated. Such a reduction would be expected to lower photopigment concentration and decrease sensitivity to light.

Why would the ectoparasitic form retain vision at all? One possibility is that the flies still use light or colour cues while moving through the host’s fur. Another is that if they are dislodged from the host, they may still need vision to find a new host on foot. A third possibility is that adult opsin expression has limited plasticity: the fly may be able to reduce overall expression, but may not have evolved a way to precisely switch off particular opsins that are no longer useful. If this last explanation is correct, L. andaluciensis carries a long-term energetic cost. The dealate ectoparasitic adult no longer needs to fly, yet it still maintains a visual system with limited utility. This may help explain why the life history of first flying to seek a host and then permanently breaking off the wings is relatively uncommon within the Hippoboscoidea.
Author: Shui-Ye You
Reference:
Santer RD et al. (2026). Visual adaptation of a biting fly that permanently foregoes flight. Journal of Experimental Biology.



Comments