
一塊來自志留紀的水蛭化石重塑蛭類起源的演化史
- Rodrigo

- 3月3日
- 讀畢需時 5 分鐘
水蛭,一種擁有黏滑身軀、以吸血行為聞名的蠕蟲,常見於溼地與潮濕的森林底層。在人類尚未書寫歷史的年代,我們的祖先就已經開始與這種難纏的環節動物打交道。
數億年來,水蛭的外型與生活方式似乎幾乎沒有改變,彷彿這種生物自誕生之初便是今日的模樣——一條柔軟、具吸盤、擅長寄生的「活化石」。但事實,真的是如此嗎?
2025 年,一塊來自美國威斯康辛州瓦基夏特異保存化石層(Waukesha Lagerstätte)的奇特化石,為這個長久以來的疑問提供了關鍵線索。這是一件出土於志留紀特列奇期(Telychian,約 4.37 億年前)的蛭類化石。這塊化石的主人被命名為志留巨蛭(Macromyzon siluricus),屬名來自希臘文,macro(巨大)+myzon(吸盤),種名siluricus則來自所屬地層志留紀。該物種體長約為51毫米,體型呈蠕蟲狀至亞披針形,前端截形,寬度向後逐漸增大,最大寬度略微位於邊緣前方。後緣寬度約為前緣寬度的兩倍。體節分節,節間呈規則的六環狀。尾部末端具大型尾吸盤。
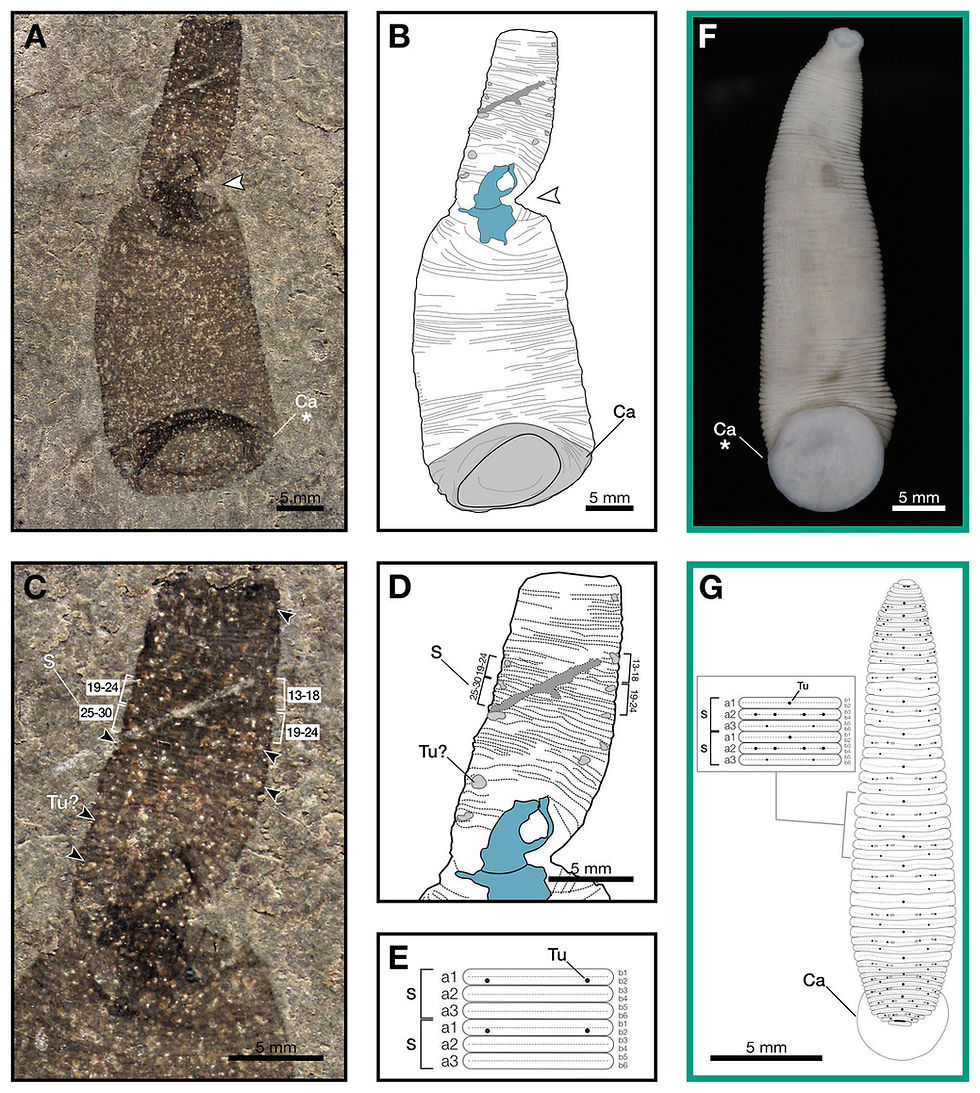
與以往所有已知的蛭類化石不同,它保存了極為清晰而完整的軟組織印痕。在過去,由於水蛭幾乎完全由軟組織構成,死亡後極易腐爛,生物學家始終難以在化石紀錄中找到牠們的「身體」。因此,研究者只能依賴兩種間接證據來推測水蛭的演化歷史。一種是依賴分子遺傳資料,透過現生水蛭與其他環節動物的 DNA 差異來回推算分化時間;另一種則是利用環節動物特有的「卵繭」化石,也就是由環帶類(Clitellata)在繁殖時分泌、用以包裹受精卵的膠質結構。

然而,這兩種證據都存在明顯限制。分子鐘的結果高度依賴模型假設,其結果容易受到突變速率變異與校準點選擇的影響。而卵繭雖然能證明「某種帶環類動物曾經存在」,卻無法告訴我們牠究竟是蚯蚓、早期水蛭,還是尚未分化完成的過渡型態,更無從得知牠的外形與生態。

正因如此,來自瓦基夏的這件化石顯得格外重要。它不僅清楚呈現出延長的體型、細密的環節結構,以及明顯的尾吸盤——這些都是現生水蛭的關鍵形態特徵——同時又保留了一些較為原始的性狀,使研究團隊得以判定:這並非現代水蛭本身,而是一種位於現生水蛭演化樹基部的「幹群水蛭」。這項發現帶來的衝擊不只在於「找到了一條古老的水蛭」,而是徹底改寫了水蛭的演化時間表。它顯示,水蛭的基本體制——包括吸盤與高度特化的體節結構——早在古生代的海洋環境中就已經出現,遠比過去根據卵繭與分子證據所推測的年代還要早上數億年。

換言之,那些我們今日在溼地與森林中見到、令人不寒而慄的吸血水蛭,並非一夕之間誕生的怪物,而是歷經漫長演化歷程、逐步從海洋走向淡水與陸地、再發展出吸血生活方式的產物。
作者: Rodrigo
參考文獻:
de Carle, D., Iwama, R. E., Wendruff, A. J., Babcock, L. E., & Nanglu, K. (2025). The first leech body fossil predates estimated hirudinidan origins by 200 million years. PeerJ, 13, e19962. https://doi.org/10.7717/peerj.19962



留言